
Introduction
Accelerated declines of terrestrial insects have been documented in many parts of the world (Ollerton 2017). Extensive spatial and temporal data from western and northern Europe, facilitated by a century-old natural history, indicates a 30-60% decline in the occurrence of common insect groups in some regions (Dirzo et al. 2014; Wagner 2020). While other long-term studies have documented similar declines across the Northern Hemisphere (as reviewed by Ollerton 2017; and Sanchez-Bayo & Wyckhuysb 2019), analogous studies and datasets for the much of the Southern Hemisphere are lacking (Pyke et al. 2023). This paucity of long-term monitoring data in the Southern Hemisphere is concerning, given the intensification of the drivers of widespread insect decline (expansion of agriculture, climate change, habitat loss/fragmentation, pesticide use, and urbanisation) (Potts et al. 2010; Dirzo et al. 2014; Sanchez-Bayo & Wyckhuysb 2019) and their potential consequences.
Insects provide fundamental ecosystem services, including biological control, nutrient cycling, and pollination, which provide inestimable environmental and economic benefits to humankind (Ollerton 2017; Loy & Brosi 2021; Wagner et al. 2021). Pollination is a crucial ecosystem service predominantly performed by insects that facilitates sexual reproduction and subsequent genetic fitness (Sands 2018; Youngsteadt & Keighron 2023) of >78% of flowering plant species (Ollerton et al. 2011). However, the ongoing loss and fragmentation of native habitats poses a key threat to many native insect pollinators and the plants they pollinate. Habitat loss and fragmentation is of high concern across urban landscapes where the connectivity and health of remaining areas of native vegetation (remnant vegetation) is essential for supporting biodiversity.
Native vegetation within Australian cities is vital for providing habitat and landscape connectivity for wildlife and is important for providing environmental enrichment for human residents (Hall et al. 2017; Ritchie et al. 2021). The resilience of native vegetation within urban environments is dependent on a diverse array of native insect pollinators which have co-evolved with native flora to maintain genetic diversity and the ecosystem values associated with successful plant reproduction (Delnevo et al. 2020a; Iwasaki & Hogendoorn 2022). Urban settings can support diverse assemblages of native insect pollinators in areas of healthy native vegetation, but the continued fragmentation of these areas to accommodate expanding urbanisation threatens this diversity through habitat and resource loss, increased isolation of insect communities (Youngsteadt & Keighron 2023), and the invasion of introduced generalist pollinators such as the European honeybee (Apis mellifera, honeybee hereafter) (Threlfall et al. 2015; Ghisbain et al. 2021).
Despite growing concerns of widespread insect declines, and the inherent values associated with pollination (Ollerton 2017; Dicks et al. 2021; Loy & Brosi 2021), the extent of insect pollinator declines and the impact on pollination services within urban Australia remains poorly understood (Pardee & Philpott 2014; Sands 2018; Ritchie et al. 2021). Furthermore, the urban pollination role of A. mellifera and subsequent impact on native pollinators is yet to be clarified with further studies needed (Elliott et al. 2021; Prendergast & Ollerton 2022; Prendergast et al. 2023). These substantial knowledge gaps can be attributed to insufficient taxonomic information, complex biotic and abiotic interactions unique to each urban environment, and the lack of long-term monitoring studies that are essential for identifying and accurately assessing the impacts of species compositional change on pollination services over time (Hall et al. 2017; Braby 2018). This not only highlights the need for fundamental research in urban pollination ecology in Australia but also the urgent requirement to establish time-series datasets that can facilitate evidence-based management actions to protect and enhance pollination services across urban habitats.
The diversity of native insect pollinators in an Australian urban habitat was assessed in a study conducted within Kings Park, Western Australia by Yates et al. (2005). At peak flowering in November 1997, Yates et al. (2005) investigated which insect visitors were frequenting the flowers of Jarrah (Eucalyptus marginata), a key mass-flowering canopy species within the remnant Kings Park bushland, at different times of the day. The study revealed that while native species richness was high, abundance was typically low for most species across the day except for the introduced honeybee which was the most common floral visitor at all times of the day. Furthermore, the relative abundance (proportion) of honeybees was found to be significantly higher than that of native bee species (Yates et al. 2005).
This current study aims to replicate the methods previously employed by Yates et al. (2005) to determine if the abundance and diversity of insect visitors and potential pollinators of Jarrah within Kings Park have changed over 26 years. This preliminary study also aims to commence the construction of a time-series dataset pertaining to Jarrah pollinator diversity by replicating the examination and analysis of insect visitor diversity across the day as per Yates et al. (2005). This will facilitate standardisation of future in-depth statistical analysis of replicated studies across multiple years and enable validity testing of the methodology in each successive study with consideration of the effects of replication and weather variables on the results. Additionally, this study aims to determine if the relative abundance of honeybees visiting Jarrah flowers compared to native bees may also have changed over 26 years. Not only can this information be used to guide further pollination research and management decisions within Kings Park, but can more broadly contribute to understanding local, national, and global changes in insect pollinator composition over time. This is of particular importance given that time-series studies pertaining to long-term insect pollinator changes in Western Australia is absent.
Methods
Study Site
Kings Park is an isolated remnant of native eucalypt and proteaceous bushland situated adjacent to the CBD within the city of Perth, Western Australia and is highly valued as a recreation, cultural, and conservation asset by the public and the State (Davis & Wilcox 2013). Perth is located on the Swan Coastal Plain bioregion in a globally recognised biodiversity hotspot with a Mediterranean climate characterised by hot, dry summers (average temperature of 32°C) and mild, wet winters (average temperature of 18°C) (Government of Western Australia 2023). Kings Park contains 267 ha of remnant native vegetation and is bound by dense urbanisation to the north and west, and the Swan River to the south and east. The bushland is dominated by Jarrah (Eucalyptus. marginata), Marri (Corymbia calophylla), Tuart (E. gomphocephala), Candlestick Banksia (Banksia attenuata), Firewood Banksia (B. menziesii), and Fraser’s Sheoak (Allocasuarina fraseriana) which have been exposed to varying degrees of disturbance including fire, introduced pests, and disease. Despite these disturbances, Kings Park remains an important conservation site with the bushland now providing vital refuge for over 320 native plants species, 80 bird species, 20 reptile species, and hundreds of native invertebrate species (Botanic Gardens and Parks Authority 2021). Management actions pertaining to the Kings Park bushland are guided by an extensive research program and a formal Tree Management Strategy by the Botanic Gardens and Parks Authority.
Jarrah (Eucalyptus marginata)
Jarrah is an important canopy species valued in the biodiversity-rich forests of Southwest Western Australia (SWWA hereafter) for providing economic and environmental resources including timber, honey, wildlife habitat, canopy cover, and firewood (McCaw et al. 2011; Bhandari et al. 2021). It is also common on the Swan Coastal Plain, particularly on soils of the Spearwood dune system, such as occurs at Kings Park. Jarrah and most other eucalypts are protandrous with a mixed mating system that facilitates a combination of outcrossing and inbreeding with no evidence of self-incompatibility (House 1997). The flowers support generalist pollination by design, with a bordering ring of stamens that inevitably come in to contact with most floral visitors, and a short distance between flowers in different gender phases. The floral structure of Jarrah is representative of most eucalypts which are visited by a range of bird, mammal, and invertebrate species, with bees, wasps, beetles, flies, and moths all documented as key insect pollinators (House 1997; Wheeler et al. 2006). Industry and government collaboration has noted generalist taxa from key pollinator groups (bird, mammal, and insect) as likely Jarrah pollinators in Western Australia (Wheen Bee Foundation 2023), although detailed species lists and observational notes specific to Jarrah are limited.
Observations of insect visitors
Detailed observations of floral visitors were documented as per the methods employed by Yates et al. (2005), with our study examining three trees over three days during peak-flowering (Table 1). Peak flowering for Jarrah in 2023 at Kings Park was mid-December compared to peak-flowering in November of 1997.
Observations were restricted to branches accessible from ground level with one c. 2 m3 section of canopy marked out per tree. The richness and abundance of insects visiting flowers were documented over two independent five-minute sampling events performed during the morning (07:00-09:00 hrs), midday (11:00-13:00 hrs), and evening (16:00-18:00 hrs) for a total of 30 minutes of observations per tree, per day. Weather conditions were documented prior to each sampling event, with ambient temperature (C°), wind speed (km/h), and rainfall (mm) recorded using the Bureau of Meteorology mobile application. Observations were made based on whether insects were probing the hypanthium disc for nectar or to the anthers of the flower. Insects were identified in the field to family or genus level where possible, or morpho-species as per Yates et al. (2005). Where required for identification purposes, a voucher specimen was obtained and identified to a minimum of family level by ARJ.
Statistical Analysis
Analysis of variance testing (with variances stabilised through transformation where required) or non-parametric equivalents (Ostertagová et al. 2014) were performed to compare the abundance and richness of insect visitors based on the time of the day and overall, for the most common orders: Hymenoptera, Coleoptera, and Diptera. Post-hoc testing (Nanda et al. 2021) was conducted to determine the statistical significance of insect visitor abundance and richness between morning, midday, and evening. A Wilcoxon signed-rank test (Harris & Hardin 2013) was conducted to compare the relative abundance (proportion) of honeybee visitors against native bee visitors at morning, midday, and evening. Mean visitor abundance and species richness (Chao & Chiu 2016) was determined with values presented as mean ± standard error (sample size).
Results
Observations of insect floral visitors December 2023
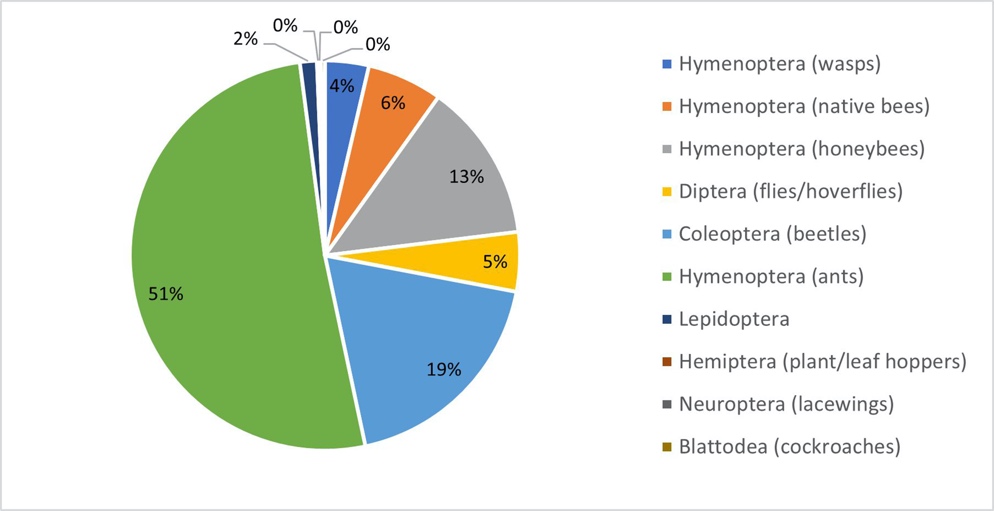
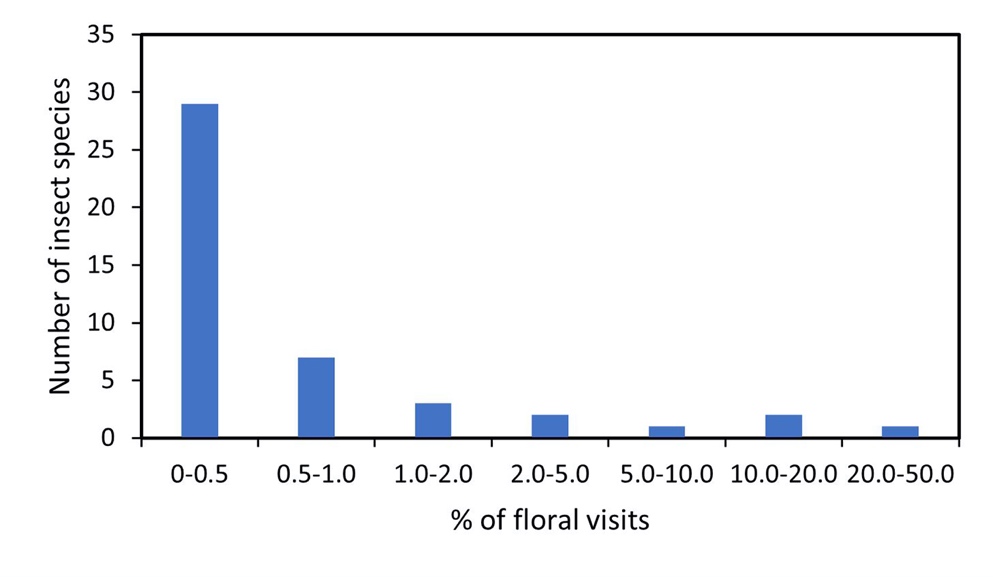
A total of 3023 observations of insects visiting the flowers of Jarrah found 45 species from 39 genera across 25 families and 7 orders (Table 2). Ants (Hymenoptera: 51% of insect visitation) and beetles (Coleoptera: 19%) were the most common floral visitor (Fig. 1), with these two groups only observed sourcing nectar. The honeybee (Hymenoptera) comprised of 13% of total floral observations followed by native bees (Hymenoptera: 6%), Diptera (5%), wasps (Hymenoptera: 4%) and Lepidoptera (2%) while the Blattodea, Hemiptera, and Neuroptera comprised of less than 0.5% of total observations each. A strong right skew was present in the distribution of insect species with 29 species recorded at or less than 0.5% of total observations, 7 species between 0.5-1%, 3 species between 1-2%, 2 species between 2-5%, 1 species between 5-10%, 2 species between 10-20%, and 1 species of ant (Formicidae gen sp.) accounting for between 20-50% of observations (Fig. 2).
_of_3023_individual_insect_visitors_(grouped_by_taxonomic_order)_to_the.jpeg)

Diurnal patterns of floral visitors
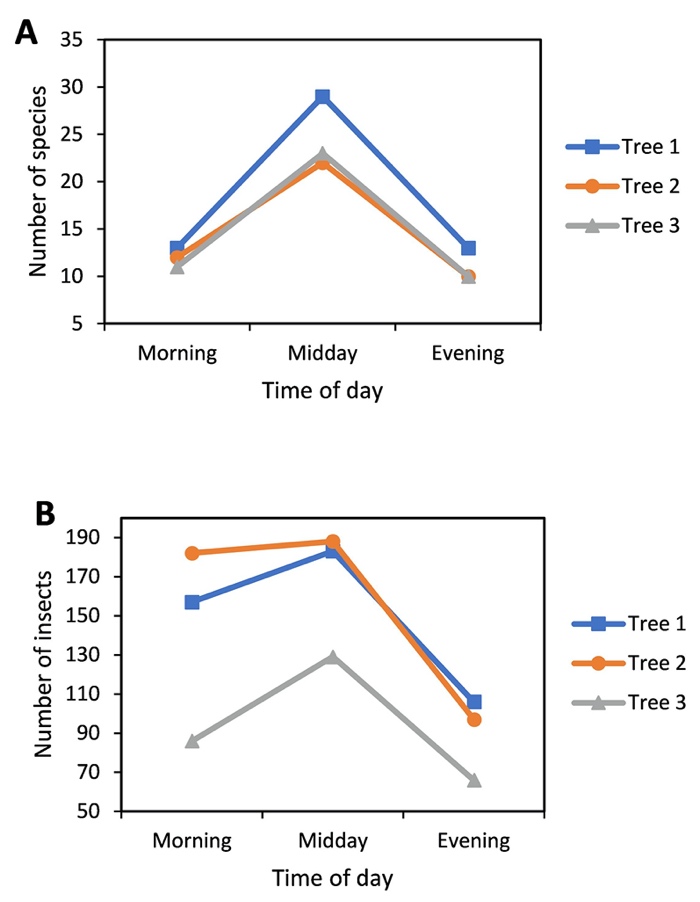
The number of species and insect abundance changed throughout the day and were both highest at midday and lowest in the evening, except for Tree 1 which recorded the similar low species richness in the morning and evening (Fig. 3). Increases in morning-to-midday abundances were less pronounced in Tree 1 and Tree 2 than in Tree 3, but all trees experienced an obvious decline in midday-to-evening abundances.
_and_number_of_individual_insects_(b)_sampled_from_t.jpeg)
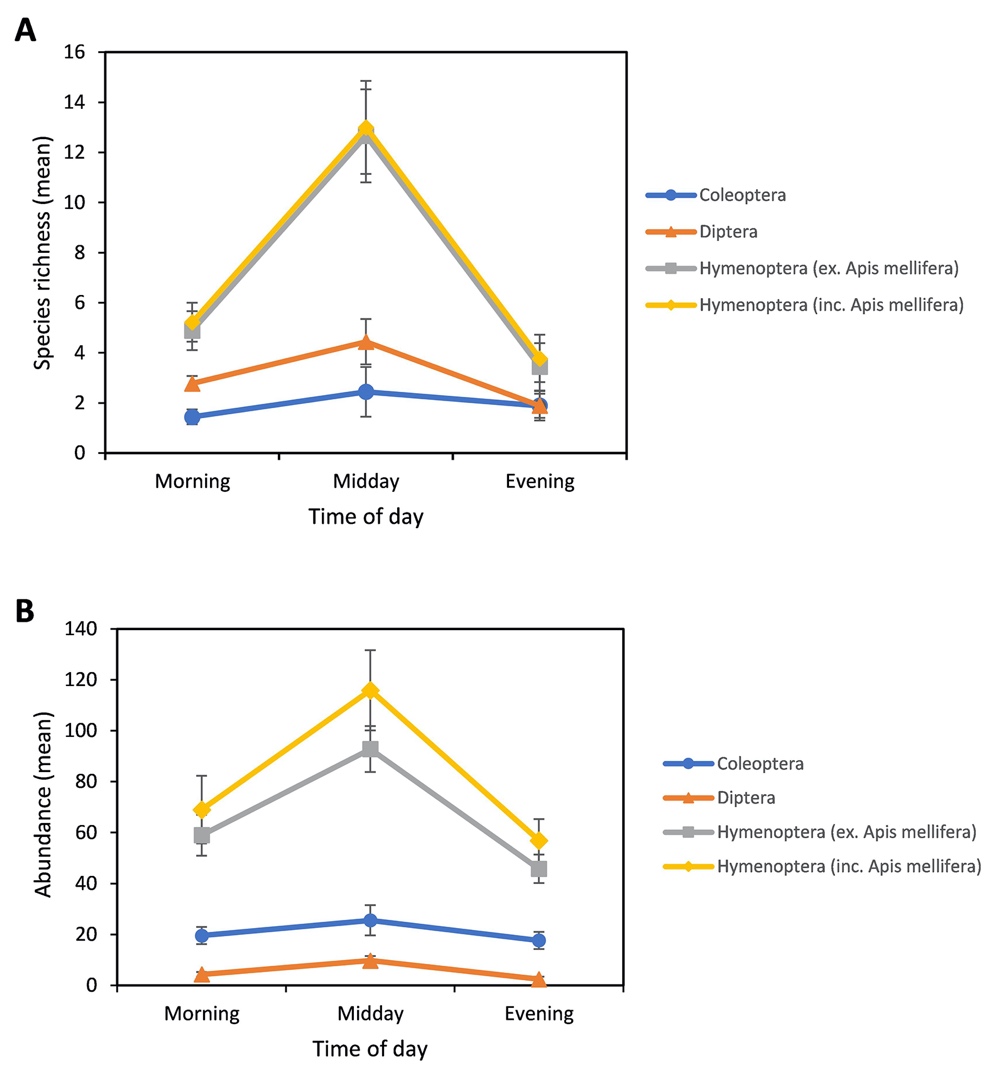
Examination of the key groups of floral visitors revealed that mean species richness across the Hymenoptera (including and excluding A. mellifera) and Diptera was highest at midday and lowest in the evening, while Coleoptera species richness was highest at midday and lowest in the morning (Fig. 4). Changes in the total species richness of both groups of Hymenoptera (including and excluding A. mellifera) across the day was significant (P = 0.005) between the evening and midday, and morning and midday, while changes in species richness of Coleoptera and Diptera across the day were not significant (Table 3).
_and_mean_abundance_(b)_of_insect_visitors_grouped_into_order.jpeg)
The mean abundance across the four keys groups of insect visitors was highest at midday and lowest in the evening, with changes across the day more prominent in both Hymenoptera groups (including and excluding A. mellifera) than in the Diptera and Coleoptera (Fig. 4). Changes in total abundance across the day for both groups of Hymenoptera was significant between the evening and midday, with the morning and midday period significant for Hymenoptera (excl. A. mellifera), and the evening and midday period significant for the Hymenoptera (incl. A. mellifera) (Table 3). Dipteran total abundance did undergo a significant change between evening and midday (Table 3), while non-parametric testing revealed that the total abundance of the Coleoptera did not change significantly throughout the day.
Ants were the most common insect visitors during the morning at 58.8%, followed by beetles (19.2%), honeybees (9.8%), native bees (4.6%), flies (4.3%), wasps (2.3%), and butterflies/moths (1%). Ants (44.3%), beetles (16.4%), honeybees (14.8%), and native bees (9.7%) were again the most abundant Jarrah visitor by proportion at midday, with all other insect groups also showing increased proportions from the morning. Cockroaches (Blattodea), leaf/plant hoppers (Hemiptera), and lacewings (Neuroptera) first occurred at midday in proportions of 0.5%, 0.3%, and 0.2%, respectively. The evening proportion of ants (55.4%), beetles (22.7%) and lacewings (0.3%) increased from midday, while honeybees (14.3%), flies (3.1%), wasps (2%), native bees (1.4%), cockroaches (0.4%), butterflies/moths (0.3%), and plant/leaf hoppers (0.1%) all decreased.
The total abundance of honeybees did display a significant change throughout the day (F = 9.3, df = 2, P = 0.015), specifically between the evening and midday (P = 0.028) and morning and midday (P = 0.019), while native bee total abundance also changed significantly (F = 59.2, df = 2, P = <0.001) between the evening and midday (P = <0.001), and morning and midday (P = <0.001). Furthermore, the relative abundance (proportion) of honeybees compared to native bees was statistically different across the day (t = -2.666, N = 9, P = 0.008). While the relative abundance of honeybees did not change significantly throughout the day, the relative abundance of native bees did express a significant change (F = 95.1, df = 2, P = <0.001) between the evening and morning (P = 0.007), the evening and midday (P = <0.001), and the morning and midday (P = <0.001).
Comparison between 1997 and 2023
A total of 46% fewer species were observed as visitors in the present study compared to 1997, while 25% of species documented in 1997 were shared across both studies. The Hymenoptera contained the most inter-study commonalities (13 shared species/morpho-species; Table 2), but also included a large native bee (Amegilla sp.) and several wasp species (Bembecinus sp., Cryptocheilus sp., Delta sp., Sphex modestus, and Polistes humilis) not documented in 1997 (Table 2; Fig. 5).
_trees_across_a_t.jpeg)
Family richness was 34% lower in the present study compared to 1997, although 10 previously unobserved families did occur with the majority of these (4 families; Crabronidae, Sphecidae, Thynnidae, and Vespidae) in the Hymenoptera (Table 2).
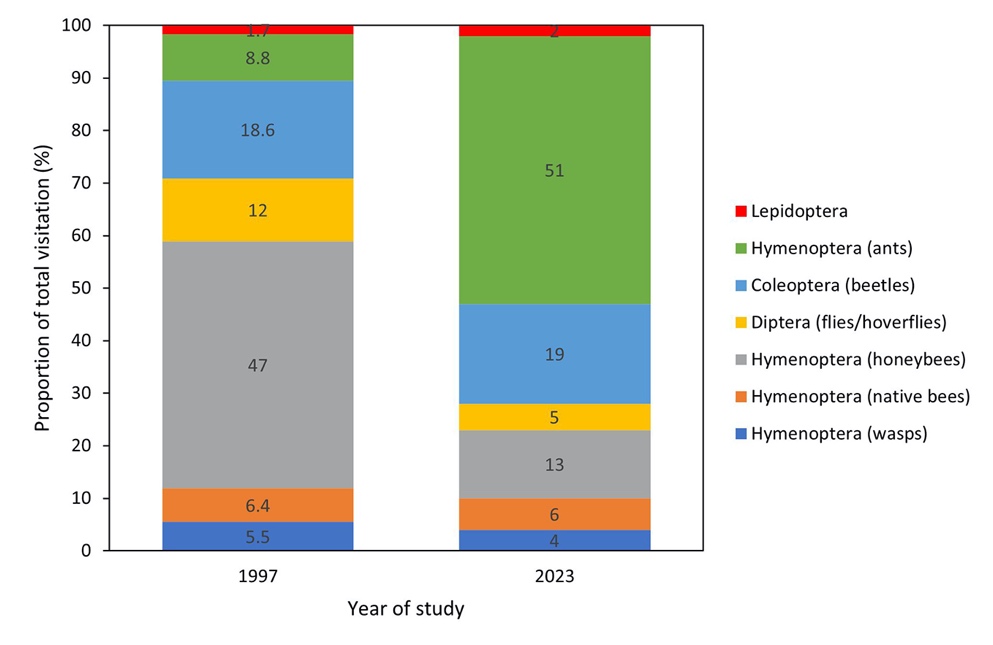
Ants were the most common visitor in the current study (51% of total observations) in contrast to honeybees which were the most common visitor 26 years ago at 47% (Fig. 6). Ants were less common in 1997 at 8.8%. Beetles (Coleoptera) were similarly common across both studies at 18.6% in 1997, and 19% in 2023, while honeybee visitation had decreased to 13% over 26 years. Flies/hoverflies (Diptera) had also decreased from 1997 to 2023 (12% to 5%, respectively). Native bees (Hymenoptera), wasps (Hymenoptera), and Lepidopterans displayed minimal change in contribution to total visitor proportions across the two studies.
_of_total_observations_contributed_by_the_most_common_insect_visitors_to.jpeg)
Discussion
Our preliminary study represents, to our knowledge, a first attempt to assess insect pollinator changes over time in Australia. We do acknowledge that tree replication in this preliminary study was limited to three trees in contrast to seven trees in 1997, albeit with a comparable temporal sampling effort between both studies. The trees sampled in our study were, in our opinion, located a substantial spatial distance apart (>0.8 km) across different Kings Park localities. However, spatial comparisons between 2023 and 1997 could not be drawn due to an absence of tree locations from 1997. Given these limitations, our results suggest that a compositional shift may have taken place in the insect taxa visiting Jarrah flowers over 26 years, with ~46% fewer species documented than Yates et al. (2005), and ants now the most prolific visitor to Jarrah flowers. Only ~25% of previously recorded visitors were documented in our study, but most species (and those recorded by Yates et al. 2005) were capable of pollinating Jarrah flowers to some extent. Members of the Formicidae (ants) however are likely an exception as they potentially impact pollen quality and viability through the excretion of antifungal compounds (Peakall et al. 1990; Lach & Thomas 2008; Takeda et al. 2021). Although ant species (both invasive and native) have been identified as nectar thieves and pollinator deterrents (Galen & Butchart 2003), ant-mediated pollination does occur, though uncommon (Das & Das 2023) it can be highly mutualistic (Delnevo et al. 2020b). While it is unlikely that ants facilitate Jarrah pollination, the impact of ant visitation on the pollination of eucalypts has not been thoroughly investigated (Lach & Thomas 2008).
In contrast to 26 years ago, ants were the most prolific visitor to Jarrah flowers in our study at all times of the day. As such, developing an understanding of how ants impact Jarrah pollen grains and the visitation rates of other insect pollinators (and pollinators in general) is an important avenue of further research. Our findings also highlight that despite a documented decline of insect pollinators in many parts of the world, some insect taxa appear less affected by human-induced stressors and may even exhibit population expansions arising from beneficial genetic traits and the decline of other species within the same habitat (Ghisbain et al. 2021). This raises additional questions pertaining to the type of impact that these ‘decline beneficiaries’ may have on critical ecosystem services like pollination.
Insect family richness was ~34% lower in our study than of that documented by Yates et al. (2005), but we did observe ten families that were not recorded as Jarrah visitors 26 years ago. Of note, was the occurrence in 2023 of members of the Hymenopteran family Vespidae in our study, with the Australian paper wasp (Polistes humilis) the only other highly eusocial wasp or bee species documented alongside the introduced honeybee. Of further note was the absence of the introduced European paper wasp (Polistes dominula) as a visitor to Jarrah flowers. While P. dominula was not documented as a visitor 26 years ago, our personal observations throughout Kings Park and other Perth metropolitan remnants have noted P. dominula as a prolific generalist visitor to many native and introduced floral species (including eucalypts) with impacts to pollination services in many other parts of the world where it has invaded (Baker & Potter 2020).
Current patterns of insect visitor abundance were similar to those documented 26 years ago but the species composition was different. We documented 3023 insect visitors to Jarrah flowers from three trees in comparison to 3278 visitors documented across seven trees by Yates et al. (2005). Akin to the 1997 study, most of our observations were dominated by a select few species, with a general low abundance recorded across all other taxa. These similar total abundances documented from a smaller sampling size (but with comparable sampling times as Yates et al. 2005) and with noticeable differences in family and species richness suggest substantial shifts in Jarrah pollinator composition at Kings Park, either through the natural fluctuations of pollinator composition over time or via human-induced stressors (Ramalho et al. 2014; Ollerton 2017). We do however caution that these data represent restricted snapshots in time, and broader conclusions about changes over time require more comprehensive and ongoing monitoring to capture natural spatial and temporal variability. Extended spatial and temporal monitoring of insect visitors to the flora within Kings Park and elsewhere should be conducted to determine the scope and drivers of pollinator fluctuations which have the capacity to impact the genetic fitness and reproductive success of native plant species. Resolving the present limitations of the study methodology, with a focus on extending tree replications beyond both existing studies, will be important to continuing this investigation of Jarrah insect diversity in the future.
The honeybee (Apis mellifera) was likely the most effective insect pollinator of Jarrah that we documented at Kings Park despite the higher abundance of ants and beetles, and the presence of solitary native species. The recognition of the honeybee as a global invasive species stems from the ability of populations to rapidly increase their invasive range and exploit floral resources through intense foraging coordination across different habitats (Cunningham et al. 2022). This rapid population expansion and eusocial behaviour has favoured the honeybee over solitary insect pollinators in urban remnants (Carmen & Jenkins 2016), but the severity of impact on native pollinators is dependent upon honeybee density, the overlap of resource requirements, and complex ecological variables unique to each remnant (Cunningham et al. 2022). In contrast to Yates et al. (2005), we found that the honeybee was not the most common visitor to Jarrah flowers due to the high abundance of ants and beetles foraging for nectar at all times of the day. However, honeybees were likely the most common effective pollinator documented visiting flowers due to their generalist morphology, dominant eusocial behaviour, the ability to quickly move between flowers while actively foraging, and the low abundance of solitary native pollinators visiting Jarrah flowers. Honeybees have been documented as a relatively effective insect pollinator of eucalypts (Moncur et al. 1995) with their larger body size capable of contacting the central stigma during foraging (Hingston & Potts 1998) but their efficacy as a pollinator of other species of Australian flora varies enormously (Paton 1996; Gilpin et al. 2019; Ayre et al. 2020).
We found that honeybees did not have exclusive access to pollen or nectar resources over native bees at any time of the day, which contrasts the findings of 26 years ago whereby native bees were absent in the morning. Our results suggest a substantial decline in the proportion of honeybee visitors to Jarrah flowers while the proportion of native bees had a minimal decline. However, while honeybee proportions were lower in our study, this relative abundance (proportion of each species sampled) was still significantly higher than native bees as per Yates et al. (2005). We did document a potential shift in the species composition of native bees with only ~37% of species in common with Yates et al. (2005), and many of the previously documented smaller bodied Hylaeus, Leioproctus and Lasioglossum species absent from our study. Of note, was the occurrence of larger bodied Amegilla native bee species in our findings that were not documented by Yates et al. (2005). While it stands to reason that smaller solitary, native bees are more likely to be outcompeted by the highly eusocial honeybee (Ollerton 2017), further studies are needed to investigate the density of honeybees at Kings Park, their impact on native insect pollinators, and the current (and emerging) pollination role they fulfil within the Kings Park bushland and across the urban remnant matrix.
At the canopy-specific level, insect biodiversity associated with Jarrah is likely driven by local determinants of site productivity such as soil fertility, habitat heterogeneity (Recher et al. 1996), and the degree of disturbance at each site (Abbott & Wills 2001). Climate conditions also strongly influence the role and composition of insect pollinators (Saunders et al. 2023). The spring rainfall averages prior to our summer study commencing were comparable to 26 years ago (473 mm and 624 mm respectively), so it is unlikely that this was a contributing factor to the differences we observed, but confounding climate effects over the long term should not be discounted. Detailed climate variables should be included in future in-depth analyses of insect visitors to Jarrah alongside improved sampling replication (spatial and temporal). While it is reasonable to expect that positive drivers of insect biodiversity within Jarrah forests will encourage diverse pollinator visitation, there have been no discernible patterns identified in how insect species are distributed across individual Jarrah trees within the same habitat (Abbott et al. 1992; Abbott & Wills 2001). Insect endemism for individual Jarrah trees was documented as rare, with most canopy visitors noted as being widespread across trees within the same habitat, but not abundant (Abbott et al. 1992; Abbott & Christensen 1994). Our study did reveal potential tree-specific insect endemism amongst solitary native bee and wasp species, but the low abundance of most recorded insect visitors highlights the need for caution when inferring potential endemism from short-term sampling events with limited replication in remnant sites. However, despite tree replication limitations, the individual trees sampled across our study were in areas exposed to the pathogen Phytophthora cinnamomi, pedestrian and vehicle traffic, and recent bushfire which may have caused some localised species isolation across individual trees. This also raises important questions for further study regarding the impacts of internal edge effects on insect pollinators that might be occurring in the Kings Park bushland near areas which act as the interface between recreation and conservation.
Conclusions
Our study suggests that the abundance of insect visitors to Jarrah flowers within Kings Park was comparable to 26 years ago, albeit with substantial reductions in family and species richness, and an evident shift in the composition of pollinator taxa. Of note was our findings that ants had replaced the honeybee as the most common visitor to Jarrah flowers, with this result raising questions about the impact of ants on the reproductive success of Jarrah and their potential influence on pollinator visitation rates. We found that no documented visitor had exclusive access to Jarrah floral resources at any time of the day, and that native bees were present at all times of the day which contrasted with their morning absence 26 years ago. Furthermore, a substantial decline in the proportion of honeybee visitors to Jarrah was evident, with minimal changes in the proportion of native bees (but with a notable shift in species composition). Despite replication limitations, our results suggest substantial changes in insect pollinator composition from 26 years ago and provide an important snapshot of current insect pollinator diversity associated with Jarrah at Kings Park which will facilitate future inter-study analyses. In our efforts to answer the ‘who’ of insect pollinators at Kings Park, we have raised important ecological questions of ‘why’ pertaining to the impact of ant species on pollination services, what drives species compositional changes in urban habitats, and the current and emerging pollination role of the introduced honeybee. These questions can only be answered through long-term monitoring and assessment of insect pollinator assemblages over time coupled with continued urban pollination research, which will not only be vital in maintaining and enhancing the ecological and social values of Kings Park, but also in addressing the decline of insect pollinators around the world.
Acknowledgments
The authors acknowledge that this work was made possible by the generous financial contributions of the Friends of Kings Park and the support of the Department of Biodiversity, Conservation and Attractions, Kings Park Science, the Botanic Gardens and Parks Authority, and Edith Cowan University.