
INTRODUCTION
Salt marshes and surrounding coastal estuarine vegetation act as dynamic intertidal ecosystems located at the interface between terrestrial and marine environments (Brearley, 2005; Reed et al., 2009). Present along most estuaries worldwide, these muddy, anaerobic and organic-rich microhabitats are characterized by halophytic vegetation – withstanding fluctuations between brackish and freshwater conditions caused by daily and seasonal change. These habitats not only are recognised for their extensive ecosystem services, serving as natural buffers against coastal erosion and flooding, but are also home to a diversity of micro- to macro-organisms, encompassing amphipods, foraminifers, bivalves and gastropods (Murray, 2014; Pennings & Bertness, 2001; Ross et al., 2009). The complex zone of rhizomes and stolons at the edge of the marsh comprises an intricate matrix, that accounts for a highly interconnected web of biological and physical processes promoting nutrient cycling and oxygen regulation (Vernberg, 1993).
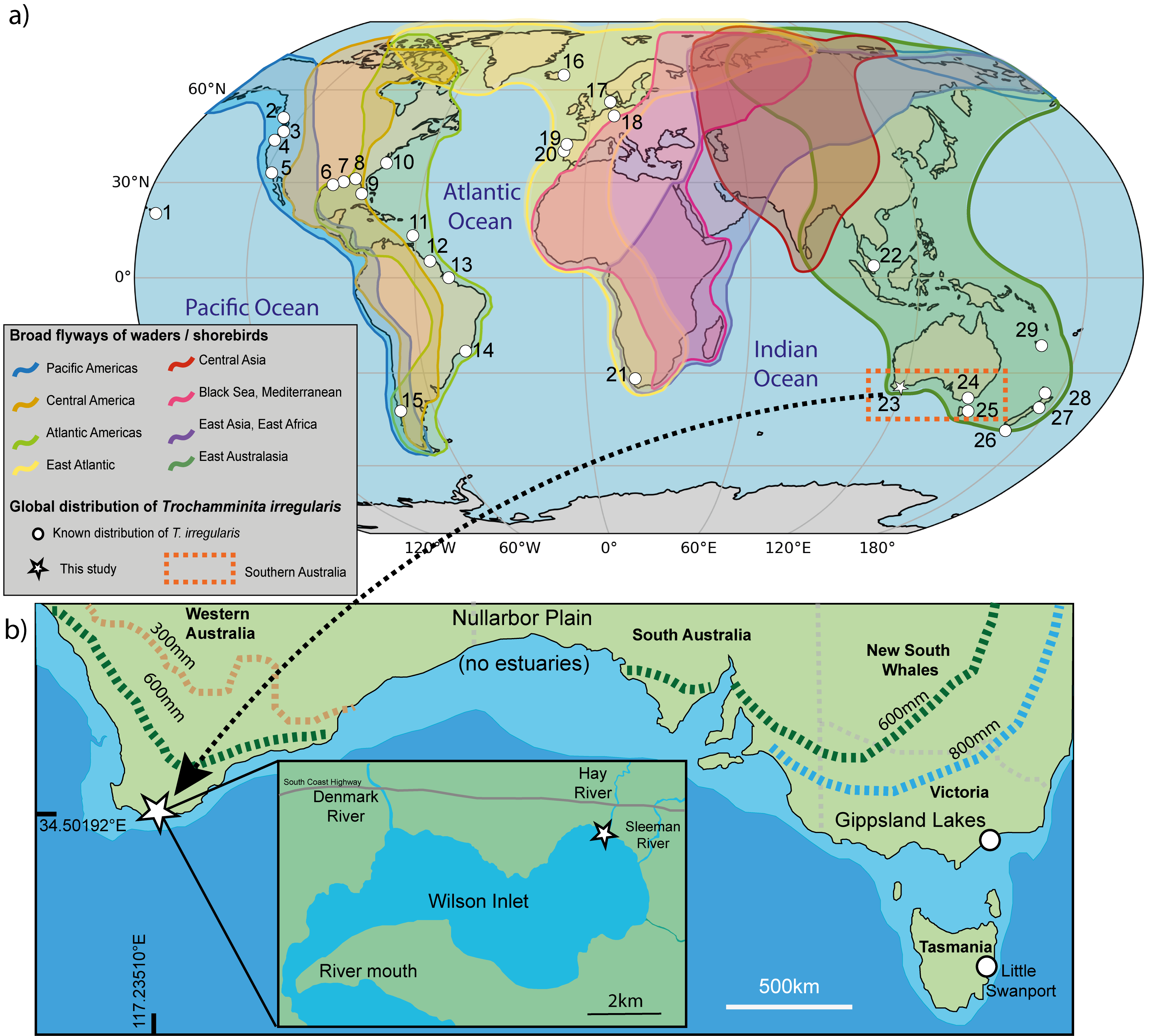
Trochamminita irregularis (Cushman and Brönnimann) is one of two species known within its genus, belonging among the organic-cemented agglutinated foraminifera that do not secrete a calcium-carbonate component in their test. These differ from carbonate-cemented agglutinated foraminifera and other groups that produce calcium carbonate through biomineralization. The species is characterised by highly variable whorl arrangements, often reaching up to four in adult specimens. Its chambers initially coil in a planispire, with the test axis shifting and twisting during later development. Trochamminita irregularis differs from the other species within its genus, Trochamminita salsa (Cushman and Brönnimann), by its asymmetrical chamber arrangement, and its wider adaptability to salinity change, compared to a more saline condition for T. salsa. Trochamminita irregularis has a broad and cosmopolitan distribution, spanning diverse geographic and ecological regions (Fig. 1, Table 1). In North America, it is found along the Atlantic and Pacific coasts, including upper marshes of Oregon (e.g., Salmon River and South Slough), Texas (Trinity River), North Carolina (Outer Banks), and Florida (Everglades). It also occurs in estuarine systems like the Pascagoula River in Mississippi and Bayou Lafourche in Louisiana. Further south, in South America, it inhabits mangrove and marsh systems in Brazil (Caeté River) and French Guiana (Kaw estuary). Its presence in Europe is notable in inland saline springs of Germany and upper marshes in Iceland and Portugal (e.g., Caminha Marsh). In the Southern Hemisphere, T. irregularis is distributed across various wetlands and marshes in Australia (e.g., Little Swanport in Tasmania) and New Zealand (e.g., Mokomoko Inlet and Pauatahanui Inlet). It has also been recorded in the saline environments of Lake Verlorenvlei, South Africa, in the estuarine lagoon of Seitu in Malaysia and in the tropical mangroves of Saint Vincent Bay, New Caledonia.
_global_distribution_(white_circles__number.png)
This preliminary study aims to (i) document the first discovery of Trochamminita irregularis in a southwestern Australian estuary, (ii) describe its micro-living habitat in Hay River, Wilson Inlet; and (iii) consider the pathways of its global distribution.
NEW LOCALITY OF TROCHAMMINITA IRREGULARIS
Wilson Inlet is a seasonally open, broad tidal inlet located on the southern coast of Western Australia (Ranasinghe & Pattiaratchi, 1999; Brearly 2005). It is a shallow estuarine basin with an area of 48 km2 and a mean depth of 1.8 m, exhibiting freshwater conditions in the winter months and brackish conditions in the summer (Lukatelich et al., 1987; Twomey & Thompson, 2001). The Hay River (Fig. 2a) is one of three main tributaries (with salinity varying from freshwater 8-10 ppt in winter to brackish 27-28 ppt in the summer months) flowing into the inlet, and it accounts for 65% of the annual streamflow into the estuary (Ranasinghe & Pattiaratchi, 1999). The Hay River catchment features two distinct landform types: with (1) a western region characterized by a rolling, dissected lateritic terrain that transitions into lateritic uplands (Lukatelich et al., 1984). In contrast, (2) the eastern portion of the catchment, which also includes the Sleeman River catchment, consists of lateritic sandplain swamps and plains interspersed with gravelly ridges (Lukatelich et al., 1984). The low relief in this area results in swampy flats surrounding the stream channels (Fig. 2b). The soils are primarily leached sands and yellow-mottled soils, which support jarrah scrub, and sandplain heaths (Lukatelich et al., 1984).
_view_of_hay_river_mouth_flowin.png)
Trochamminita irregularis is found in low abundance in shore-line environments near the confluence of Hay River and Wilson Inlet at coordinates 34.970089°S, 117.4692702°E (Fig. 2c). It forms <20% of the marsh foraminiferal fauna in a systematic pick of dead and live individuals, alongside Ammonia haigi Haywood and Holzmann, Trochammina inflata (Montagu), and Elphidium sp., and lives among riparian vegetation including Sarcocornia quinqueflora Bunge ex Ung.-Sternb. (Fig. 2d), Baumea sp. and Juncus sp. (Fig. 2e). The substrate is composed of fine silty mud with very rare shelly material.
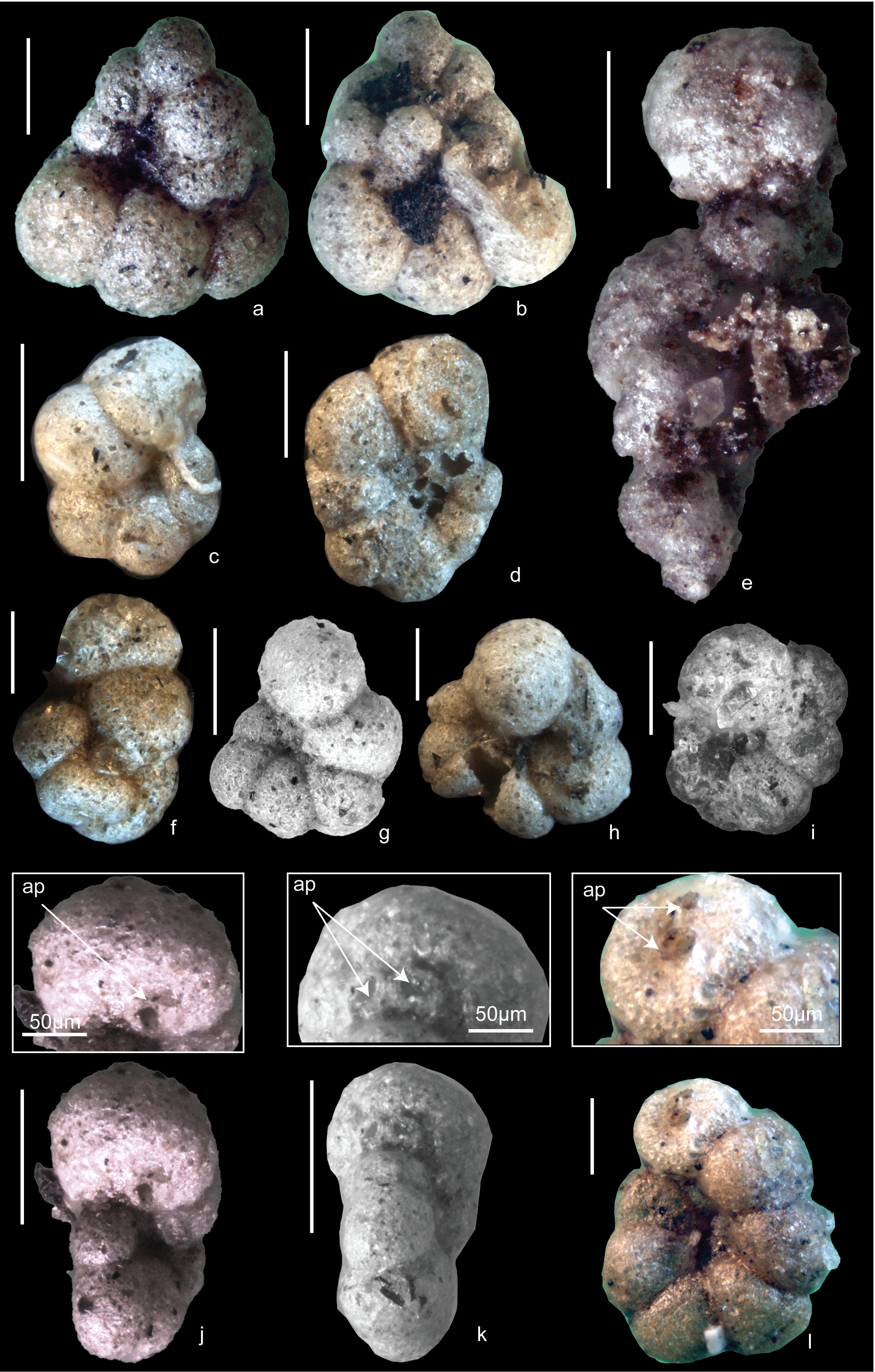
Morphological variability (Fig. 3a–l) of the test of T. irregularis could likely be shaped by the intertwining pattern of above-ground stolons at the base of the vegetation. The multiple apertures and their varying position either at the base of the ultimate chamber or areal in the apertural face of this chamber (Figs. 3j, k, l) may be a potential morphological adaptation to maximise food uptake and to facilitate the protrusion of the pseudopodia in 3-dimensions, while attached to the filamentous stolons/rhizomes. In the slightly deeper part of the Hay River (>0.5 m water depth), the seagrass Ruppia megacarpa Manson is found in occasional patches growing in a mix of sandy and silty mud. Trochamminita irregularis was not found in this habitat.
_from_the_hay_river_sites_shown_in__fi.png)
DISCUSSION
Associated foraminifers with Trochamminita irregularis
Worldwide species associations
Trochamminita irregularis commonly coexists with a variety of foraminiferal species that inhabit similar marsh, estuarine, and mangrove ecosystems. In temperate and subtropical marshes, it is often found alongside species such as Entzia macrescens (Brady), often referred to Jadammina, and Miliammina fusca (Brady), which are well-known indicators of high-marsh environments (Table 1). The association with Trochammina inflata, another marsh specialist, is consistent in regions like Oregon, New Zealand, and South America. In saline springs, it is found with E. macrescens and Siphotrochammina lobata Saunders, highlighting its adaptability to high salinity and fluctuating environmental conditions. In tropical and subtropical ecosystems, T. irregularis cohabits with Arenoparrella mexicana (Kornfeld), Haplophragmoides wilberti Anderson, and species of Ammonia (Table 1). Other notable cohabitants include Balticammina pseudomacrescens Brönnimann, Lutze & Whittaker in northern marshes (e.g., Oregon) and Trochamminita salsa, a species found in South America, Germany, and New Zealand. In Portugal, Australia, and Malaysia, T. irregularis is also associated with species often referred to as Quinqueloculina “seminula” (Linnaeus), Paratrochammina haynesi (Atkinson), and Polysaccammina ipohalina Scott, which are indicative of brackish to marine transitions. By understanding these co-occurrences, it does underline the ecological versatility and habitat diversity in which T. irregularis evolved but also raises significant questions about dispersal of the species.
Hay River species association
Based on previous and continuing studies on Western Australian estuaries, the Hay River site described here is the only locality where Trochamminita irregularis has been found in the State (Table 2). The dominant foraminiferal species at the Hay River study site are the calcareous trochospiral Ammonia haigi and the organic-cemented agglutinated trochospiral Trochammina inflata. Apart from the Hay River site, T. inflata is known from 12 of the other 28 world-wide T. irregularis sites listed on Table 1, and species of Ammonia were recorded from six of the sites. These species are known from many of the other estuaries in south-west Western Australia. Questions exist, that require much further work and cannot be answered in this short note, about why different species are associated with Trochamminita in the marsh associations at different localities (Table 1), and why T. irregularis has not been found at other sites in Western Australia containing associated marsh foraminifers (based on extensive unpublished work on most estuaries from Esperance in the east to Carnarvon in the north-west).
Biogeographical fragmentation: potential means of distribution
The global distribution of T. irregularis is highlighted by a fragmented pattern ranging from tropical to temperature regions, and by differences in associated species almost all of which are confined to marshes and river-margin situations under mainly brackish water conditions. Most estuaries are disconnected spatially and in geological time. This is very apparent between the two known occurrences of Trochamminita irregularis on the southern coastline of Australia, Gippsland Lakes in the east and Hay River in the west, with about 1500 km of the >2500 km shoreline between these sites lacking river inflow and estuaries.
Dispersal of estuarine foraminifera has been suggested to occur by different means including anthropogenic interventions (ballast water, e.g., Tremblin et al., 2021), or from natural mechanisms (drifting of foraminiferal propagules in the current, e.g. Alve, 1999, and rafting e.g., Finger, 2018). Bird-mediated dispersal of riverine invertebrates including foraminifera has been suggested in the literature through ectozoochory and endozoochory, where the biota get into feathers, feet or directly in the birds digestive systems (Coughlan et al., 2017; Riedel et al., 2011). Dispersal events by birds may be rare but because of the large migratory waterbird populations and the countless days of migration from their departure point to their destination, the cumulative rate of rare dispersals make these significant.
In the Australian context, dispersal of estuary-bound foraminifers must have been across large terrestrial spaces (either north-south or east-west) or over large tracts of ocean surrounding the continent. Australia is located at the edge of one of eight major flyways for migratory land and waterbirds (Fig. 1). This flyway accounts for about 700 species of birds flying from as far away as Siberia in the north and to New Zealand in the South. In their foraging, waterbirds may unintentionally transport T. irregularis through their feathers, feet, or digestive systems as they migrate across vast distances. Five species of waterbirds including the Eurasian Coot (Fulica atra), Red-necked Stint (Calidris ruficollis), Silver Gull (Chroicocephalus novaehollandiae), Fairy Tern (Sternula nereis) and Caspian Tern (Hydroprogne caspia), share the three localities where Trochamminita irregularis has been recorded in Australia (Birdlife Western Australia, 2024; Bryant, 2002; Hansen et al., 2024). Under extreme conditions foraminiferal protoplasm can remain viable for long periods of time, up to two years, as described by Alve and Goldstein (2010). It has also been shown from fish-gut contents that more than 20% of foraminiferal protoplasm is still viable even after days of being digested (Guy-Haim et al., 2017). In the case of T. irregularis, their asexual reproduction, as well as their flexible, imperforate, and highly variable tests built on an organic template, and their ability to construct their tests using surrounding fine siliceous sand grains, probably make them likely to quickly regenerate a population when an environmentally suitable habitat is encountered. The other taxa associated with T. irregularis could have been transported via the same mechanisms. The occurrence of T. irregularis in Hay River adds to the presently known record of the species distribution and emphasizes the ecological importance of studying salt-marsh environments in Western Australia and elsewhere and using foraminifers as markers for not only possible recent introductions, but also as environmental proxies.
Future work should involve (1) genetic analysis to understand the connectivity of the various estuarine Trochamminita populations worldwide including in areas of overlap of the migratory flyways, as well as the genetic drift along possible migratory pathways; (2) seasonal ecological monitoring of T. irregularis in the Hay River, to note species dynamics under varying climatic conditions; and (3) analyses of mud detritus attached to feathers and feet as well as the faecal content of migratory birds to determine if attached mud and faeces could facilitate the foraminiferal migration.
CONCLUSIONS
Trochamminita irregularis, a species recognized for its global distribution, previously recorded in Australia only in the Gippsland Lakes (Victoria) and Little Swanport Estuary in Tasmania, has now been identified in the Hay River of Wilson Inlet, located in south-western Australia. This species is distinguished by its variable chamber morphology, multiple apertures, and flexible test. It occupies the muddy substrate at the base of upper marsh vegetation, in a habitat where environmental factors such as tidal changes and salinity fluctuations within a general hyposaline range may shape its development. The fragmentary wide distribution of the species may be associated with migratory waterbirds, which could facilitate its spread in mud attached to feathers and feet or in faeces. Further molecular and ecological research is essential to clarify its dispersal mechanisms and ecological role, particularly as an important indicator species in coastal ecosystems.
ACKNOWLEDGEMENTS
This paper is the result of discovering Trochamminita irregularis in samples collected in September 2019 by David Haig, Bruce Hayward, Mark Gunson and Jenny Bevan, in the Hay River. David Haig is thanked for his assistance in the field and in discussions; Siri Kellner of the University of Western Australia Earth Science Museum is thanked for her curatorial assistance. The authors are grateful to the two anonymous reviewers for their helpful comments and suggestions that have improved the manuscript. All materials are curated in the Earth Science Museum of the University of Western Australia.