
_in_western_australia.tiff)


INTRODUCTION
Bryozoans are among the dominant macrofossils in the Permian marine sediments of Australia (e.g., Teichert 1951; Wass 1969; Engel 1987; Engel and Ross 1993; Reid and James 2008, 2010). Extensive bryozoan faunas from the Permian of Eastern and Western Australia as well as Tasmania were studied by many authors (e.g., Nicholson and Etheridge 1886; Bretnall 1926; Crockford 1957; Ross 1963; Wass 1968; Reid 2003). In Western Australia bryozoans are especially rich and abundant in limestone facies of the Lower Permian Callytharra Formation (Fig. 1; Sakmarian–Artinskian; Haig et al. 2014) Outcrops of the formation in the Merlinleigh Basin (part of the broader Carnarvon Basin of Hocking et al. 1987, and Southern Carnarvon Basin of Hocking 1994) contain highly diverse bryozoans. However, only few publications are devoted to their study (e.g., Hudlestone 1883; Hinde 1890; Bretnall 1926; Crockford 1944a, b; Key et al. 2025). An overview of the bryozoan fauna from the Callytharra Formation was given by Engel and Ross (1993, p. 95). Their list includes 29 species. However, a study of unpublished material available to the present author indicates the presence of about 50 bryozoan species in the formation.
This paper aims to diagnose, describe and illustrate three new species belonging to the Order Trepostomata from three localities of the Callytharra Formation in the Merlinleigh Basin. This will be a prelude to a comprehensive study of the bryozoan fauna from the formation.
Geological setting
Permian strata of Western Australia were deposited within the East Gondwana interior rift system (Haig et al. 2014) which represents a broad N-S zone of late Paleozoic and early Mesozoic interior-rift and interior-sag basins in East Gondwana that during the Jurassic-Cretaceous developed into the western margin of the Australian continent (Stagg and Colwell 1994; Langhi and Borel 2005). The Callytharra Formation was deposited above glacially influenced sediments in a narrow rift basin (Merlinleigh Basin) far from the open ocean (Haig et al. 2014, their fig. 2; 2017, their figs. 1, 2).

The Callytharra Formation consists of a series of shallowing upward cycles with an overall progradational pattern and is up to 125 m thick in outcrop (Haig et al. 2014, their figs. 14–18). Each parasequence consists of a unit of mudstone, sandy mudstone or muddy sandstone at the base with a thin unit (usually < 10 cm thick) of skeletal carbonate-cemented mudstone, packstone or grainstone at the top. At the top of each parasequence set, amalgamated limestone beds (up to 0.75 m thick) are present. At least five parasequence sets are recognized in the formation. The limestones of the Callytharra Formation represent bryozoan-rich carbonates that have a very rich marine benthic invertebrate fauna dominated by bryozoans, brachiopods and crinoids. As shown by Haig et al. (2014), Tubiphytes (of suspected algal affinity), and smaller foraminifers belonging to the Fusulinata are present in the muddy carbonates of the lower Callytharra Formation and these suggest at least temperate, if not warmer, water conditions for the upper Sakmarian part of the formation (Haig et al. 2014, 2018).
MATERIAL AND METHODS
All the studied material comes from three localities of the Callytharra Formation in the Southern Carnarvon Basin, Western Australia (Figure 2; Table 1). The age of samples follows Haig et al. (2014, their fig. 13) based on the occurrences of mainly ammonoids as explained in their appendix B.
_in_western_australia.tiff)
From the studied samples thin sections were made (24x48 mm and 50x50 mm). Bryozoan morphology has been studied using a binocular microscope. Morphologic character terminology is partly adopted from Boardman (1960) and Anstey and Perry (1970). The following morphologic characters were measured and used for statistics in the studied material: Branch width, branch thickness, exozone and endozone width, axial ratio (ratio of endozone width to the branch width), autozooecial aperture width, aperture spacing (along branch, diagonally), acanthostyle diameter, exilazooecia width, wall thickness in exozone.
For branched bryozoans, the Bryozoan Skeletal Index (BSI) has been calculated, using the formula (Exozone Width x Exozonal Wall Width/Aperture Width) x 100 (Wyse Jackson et al. 2020). The spacing of structures is measured as a distance between their centres. Statistics were summarized using arithmetic mean, sample standard deviation, coefficient of variation, and minimum and maximum values. The studied thin sections are deposited at the Western Australian Museum, Perth.
Taxonomy
Phylum Bryozoa Ehrenberg 1831
Class Stenolaemata Borg 1926
Superorder Palaeostomata Ma, Buttler and Taylor 2014
Order Trepostomata Ulrich 1882
Suborder Amplexoporina Astrova 1965
Family Stenoporidae Waagen and Wentzel 1886
Genus Nikiforopora Dunaeva 1964
Type species
Batostomella concentrica Nikiforova 1927. Beshevo Limestones (Mississippian, Serpukhovian); Beshevo village, left shore of Kalmius river, Donbass, Donetsk region, Ukraine.
Diagnosis
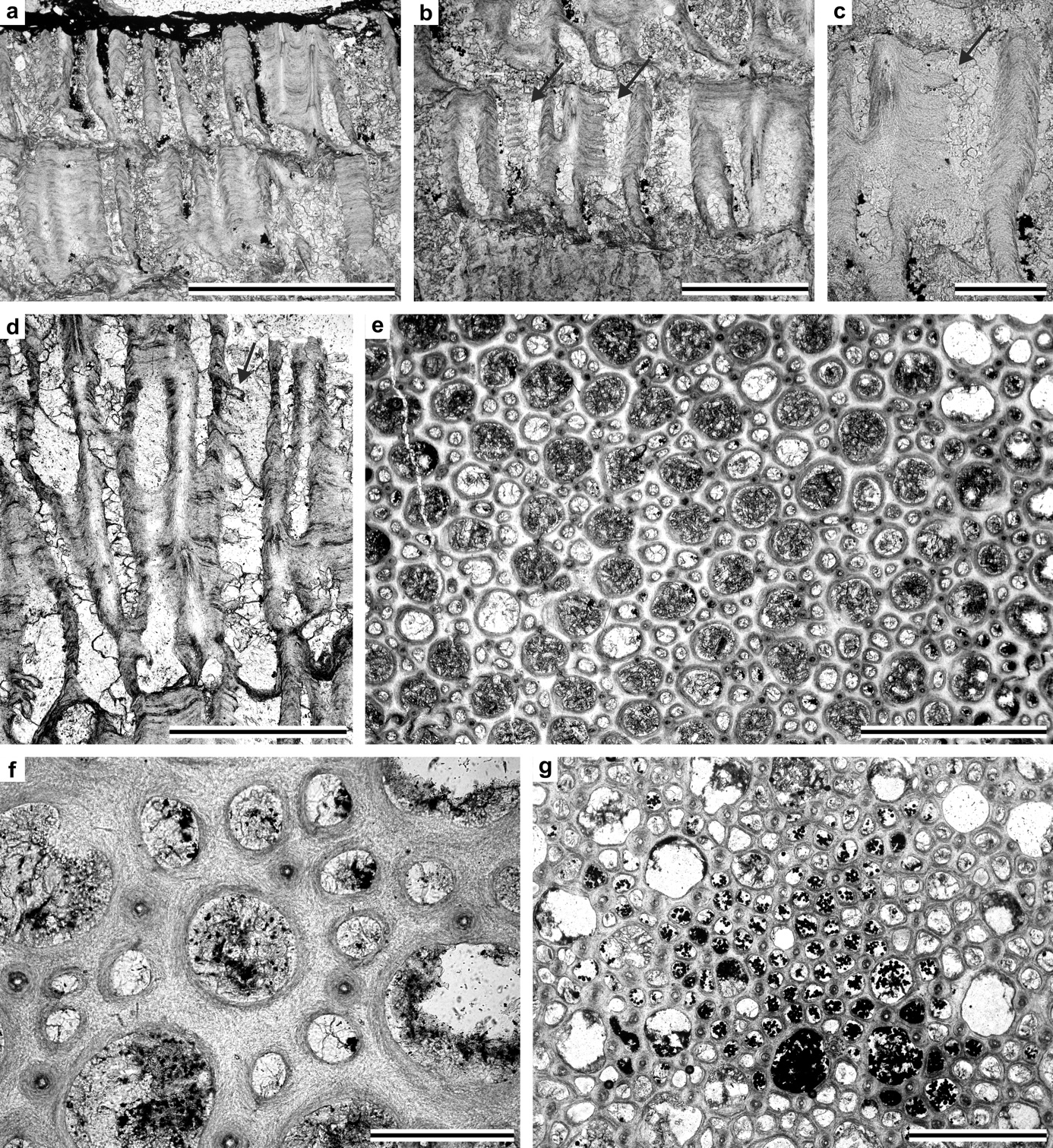
Ramose or encrusting colonies. Autozooecia polygonal in transverse section of endozone, bending abruptly in exozone and intersecting colony surface at high angles. Autozooecial apertures rounded to sub-polygonal. Exilazooecia small, circular in transverse section, not common. Diaphragms thin, rare. Ring septa few, thickened at boundary of endozone and exozone; with proximal side thicker than distal. Heterostyles (cortex spherules) abundant in exozone, roughly arranged along zooecial boundaries, forming circular-polygonal rings around the autozooecia. Exozonal styles large, located on zooecial boundaries. Walls thin and smoothly curving in endozones; regularly thickened, laminated, merged in exozone; cingulum absent. Maculae unknown.
Remarks
Nikiforopora Dunaeva 1964 differs from Stenopora Lonsdale 1844 and Stenodiscus Crockford 1945 in the development of ring septa and the presence of heterostyles in the exozone. Nikiforopora differs from Tabulipora Young 1883 in presence of the heterostyles in the exozone.
Occurrence and geological age
Upper Devonian (Famennian) – Lower Permian (Artinskian); Eurasia, Australia.
Nikiforopora haigi sp. nov.
Etymology
The new species is named after David Haig (Perth), to acknowledge of his great help during the collection of the Permian bryozoans in Western Australia.

Holotype
2025.145a-f (seven thin sections of a single colony).
Paratypes
2025.146a-g, 2025.147a-g, 2025.148a-d, 2025.149a-b, 2025.150a-c, 2025.151a-b, 2025.152a-c, 2025.153.
Type stratum
Callytharra Formation, Lower Permian (Holotype and paratypes 2025.146a-g, 2025.147a-g, 2025.148a-d from the lower Artinskian Jimba Jimba Calcarenite Member of the Callytharra Formation; other paratypes from lower Artinskian, upper Callytharra Formation equivalent to the Jimba Jimba Calcarenite Member).
Type locality
Jimba-Jimba Calcarenite type section, Merlinleigh Basin, Western Australia.
Other occurrences
Upper Callytharra Formation (equivalent to the Jimba Jimba Calcarenite Member), Lower Permian (lower Artinskian); Dead Man’s Gully section and Blair’s Camp section, Merlinleigh Basin, Western Australia.
Diagnosis
Branched colonies with distinct endozones, secondary overgrowth occurring; autozooecial diaphragms rare, ring septa abundant; acanthostyles large, 2–5 surrounding each autozooecial aperture, locally absent; heterostyles (cortex spherules) large, 7–15 arranged in a single row between acanthostyles; exilazooecia rare to common, small and short.
Description
Branched colonies with distinct endozones, 3.8–5.9 mm in diameter, with 0.5–1.4 mm wide exozones and 2.1–4.1 mm wide endozones. Axial ratio is 0.53–0.80. BSI is 69.2. Secondary overgrowth occurring, 0.6–1.3 mm thick. Autozooecia long in endozones bending abruptly in exozones and intersecting colony surface at high angles, having polygonal shape in transverse section of endozone, containing rare diaphragms and abundant ring septa. Ring septa originating in the late endozone, common throughout the exozone, with inner rims curved proximally. Ring septa in the transition between endo- and exozones often thickened at their proximal parts. Autozooecial apertures rounded-polygonal. Acanthostyles large, having narrow hyaline cores and wide laminated sheaths, 2–5 surrounding each autozooecial aperture. Locally acanthostyles absent. Heterostyles (cortex spherules) large, having narrow laminated sheaths and wide irregularly wrinkled cores, arranged in a single row between acanthostyles, 7–15 surrounding each autozooecial aperture. Exilazooecia rare to common, small and short, rounded to oval in transverse section. Endozonal walls laminated with dark boundaries, 0.005–0.015 mm thick; exozonal walls thickened, laminated, merged, without distinct autozooecial boundaries, 0.07–0.25 mm thick. Maculae not observed.
Remarks
Nikiforopora haigi sp. nov. is similar to the N. aliutovensis (Schulga-Nesterenko 1955) from the Pennsylvanian (Moscovian) of Russia but differs from the latter in larger autozooecial apertures (aperture width 0.12–0.30 mm vs. 0.07–0.10 mm in N. aliutovensis). Nikiforopora haigi sp. nov. differs from N. concentrica (Nikiforova 1927) from the Mississippian (Serpukhovian) of Ukraine in larger autozooecial apertures (aperture width 0.12–0.30 mm vs. 0.16–0.21 mm in N. concentrica) as well as in larger acanthostyles (acanthostyle diameter 0.04–0.13 mm vs. 0.06–0.07 mm in N. concentrica).
Family Dyscritellidae Dunaeva & Morozova 1967
Genus Iraidina Kiseleva 1982
Type species
Iraidina damperovi Kiseleva 1982. Vladivostok Horizon, Upper Permian (Roadian–Wordian); Primor’ye, Russia.
Diagnosis (emended)
Encrusting and ramose colonies. often with secondary overgrowth. Autozooecia long in endozones and short recumbent in encrusting colonies, with irregular polygonal shape in transverse section. Autozooecial apertures rounded to sub-polygonal. Diaphragms thin, uncommon. Exilazooecia few to common, varying in size, circular to irregular in transverse section. Large acanthostyles present. Autozooecial walls indistinctly laminated in endozone; laminated, merged in exozone; cingulum absent. Thick mural projections, some straight, others hook-shaped in longitudinal section, closely and irregularly spaced from zooecial wall within autozooecial chamber. Maculae composed of exilazooecia and acanthostyles present.
Remarks
Iraidina Kiseleva 1982 is similar to Dyscritella Girty 1911 but differs from it in presence of mural spines, diaphragms, and acanthostyles of only one size. Iraidina differs from Stenoporella Bassler 1936 in its shape of mural spines (hook-shaped vs. rounded, blunt spines in the latter genus), abundant exilazooecia and absence of ring septa.
Occurrence and geological age
Three species were previously known: Iraidina damperovi Kiseleva 1982 and I. ramosa Lisitsyn 1986 from the Upper Permian of Russia, I. maxima Morozova 1991 from the Upper Permian of Mongolia. Iraidina dendroidea sp. nov. and I. multicava sp. nov. are described from the Callytharra Formation of Western Australia (present paper).
Iraidina dendroidea sp. nov.
Etymology
The species name refers to the branched colony form (from Greek “dendroeidés” = dendroid).
Holotype
2025.154a-k (eleven thin sections of a single colony).
Paratypes
2025.155, 2025.156a-c, 2025.157a-e, 2025.158, 2025.159, 2025.160, 2025.161a-m
Type stratum
Upper Callytharra Formation (equivalent to the Jimba Jimba Calcarenite Member), Lower Permian (lower Artinskian)
Type locality
Dead Man’s Gully section, Merlinleigh Basin, Western Australia.

Other occurrences
Section A, 21.5 m above base of Callytharra Formation, Lower Permian (Sakmarian); Blair’s Camp section, Merlinleigh Basin, Western Australia (paratype 2025.160). Section B, 39 m below top of Callytharra section (? Artinskian), probably in lower part of Jimba Jimba Calcarenite Member equivalent (above a level with sandstone beds); Blair’s Camp section, Merlinleigh Basin, Western Australia (paratype 2025.161a-m).
Diagnosis
Ramose branched colonies; exozones distinct; secondary overgrowth occurring; 4–9 exilazooecia and 1–6 acanthostyles surrounding each autozooecial aperture; mural projections present; autozooecial diaphragms absent; maculae consisting of exilazooecia present.
Description
Ramose branched colonies, 2.9–6.6 mm in diameter, with 0.4–2.3 mm wide exozones and 1.3–5.6 mm wide endozones. Axial ratio is 0.21–0.77. BSI is 74.3. Secondary overgrowth occurring, 1.50–2.25 mm thick. Autozooecia long in endozones bending abruptly in exozones and intersecting colony surface at high angles. Autozooecial apertures rounded, often petaloid due to indenting acanthostyles. Exilazooecia restricted to exozone, rounded-polygonal in transverse section, abundant, often separating autozooecia, 4–9 surrounding each autozooecial aperture. Acanthostyles large, abundant, 1–6 surrounding each aperture, having distinct calcite cores and dark laminated sheaths. Autozooecial walls indistinctly laminated, 0.005–0.010 mm thick in endozones; laminated, merged, 0.05–0.14 mm thick in exozones. Mural projections, straight to curved proximally, originating from the proximal wall of the zooecial chamber, moderately spaced. Rare maculae consisting of exilazooecia and acanthostyles present, 0.59–0.61 mm in diameter.
Remarks
Iraidina dendroidea sp. nov. differs from I. damperovi Kiseleva 1982 in having smaller autozooecial apertures (aperture width 0.10–0.20 mm vs. 0.20–0.27 mm in I. damperovi). Iraidina dendroidea differs from I. maxima Morozova 1991 in having smaller branches (branch diameter 2.9–6.6 mm vs 9.5–10 mm in I. maxima) as well as in having smaller apertures (aperture width 0.10–0.20 mm vs. 0.20–0.30 mm in I. maxima). Iraidina dendroidea differs from I. ramosa Lisitsyn 1986 in having smaller autozooecial apertures (aperture width 0.10–0.20 mm vs. 0.18–0.28 mm in I. damperovi).
Iraidina multicava sp. nov.
Etymology
The species name refers to the abundant exilazooecia of the new species (from Latin “multicavum” – porous).
Holotype
2025.162a-g (seven thin sections of a single colony).
Paratypes
2025.162a-g, 2025.163a-g, 2025.164a-e, 2025.165, 2025.166a-b, 2025.167, 2025.168a-e.
Type stratum
Callytharra Formation, Jimba Jimba Calcarenite Member, Lower Permian (lower Artinskian).
Type locality
Jimba-Jimba type section, Southern Carnarvon Basin, Western Australia.

Other occurrences
Upper Callytharra Formation, Jimba Jimba Calcarenite Member equivalent (lower Artinskian); Dead Man’s Gully section, Merlinleigh Basin, Western Australia.
Diagnosis
Encrusting colonies, occasionally multilaminar; 5–12 exilazooecia and 2–6 acanthostyles surrounding each autozooecial aperture; mural projections present; autozooecial diaphragms absent; maculae consisting of exilazooecia present.
Description
Encrusting and submassive colonies, occasionally multilaminar. Separate sheets 0.3–1.0 mm thick. Multilamellar colonies 1.1–14.0 mm in thickness. Epitheca 0.015–0.018 mm thick. Autozooecia short in endozones bending abruptly in exozones and intersecting colony surface at high angles. Autozooecial apertures rounded, often petaloid due to indenting acanthostyles. Exilazooecia polygonal in transverse section, abundant, often separating autozooecia, 5–12 surrounding each autozooecial aperture. Acanthostyles large, abundant, 2–6 surrounding each aperture, having distinct calcite cores and dark, laminated sheaths. Autozooecial walls indistinctly laminated, 0.005–0.015 mm thick in endozones; laminated, merged, 0.04–0.09 mm thick in exozones. Mural projections, straight to curved proximally, originating from the proximal wall of the zooecial chamber, moderately spaced. Maculae consisting of exilazooecia and acanthostyles present, 0.4–0.8 mm in diameter.
Remarks
Iraidina multicava sp. nov. differs from I. damperovi Kiseleva 1982 in smaller autozooecial apertures (aperture width 0.12–0.23 mm vs. 0.20–0.27 mm in I. damperovi). Iraidina multicava differs from I. dendroidea sp. nov. in having larger autozooecial apertures (average aperture width 0.18 mm vs. 0.14 mm in I. dendroidea) and in more abundant exilazooecia (average number of exilazooecia per autozooecial aperture 9.0 vs. 5.8 in I. dendroidea).
DISCUSSION
Trepostome bryozoans represent a significant part of the Lower Permian bryozoan assemblages in Australia. However, our knowledge about them is still relatively poor. In the present paper three new trepostome species from the Callytharra Formation of the Southern Carnarvon Basin are described: Nikiforopora haigi sp. nov., Iraidina dendroidea sp. nov., and I. multicava sp. nov. The genera Nikiforopora and Iraidina are reported from the Permian deposits of Australia for the first time. Nikiforopora is known from the Upper Devonian (Famennian) to Pennsylvanian (Moscovian). All known species were described from Russia, Ukraine, Kazakhstan and Mongolia. The new species Nikiforopora haigi is the youngest representative of the genus.
In contrast, the genus Iraidina was previously known from the Upper Permian (Roadian) of Urals and Far East of Russia as well as Mongolia. The two new species are the oldest representatives of the genus.
It appears possible, however, that these genera may be present in other Permian formations of Australia as well as in contemporary deposits of adjacent territories. They could be misidentified because of misinterpretation of their morphology. Nikiforopora is similar to Tabulipora, but differs from the latter in the kind of styles and in the wall structure. These characters can only be studied in oriented thin sections under high magnification. The same problem concerns Iraidina which differs from Dyscritella in presence of mural projections in their autozooecial walls. This morphology cannot be seen without thin sections. Unfortunately, a significant number of older publications describe bryozoans without the use of thin sections. Re-study of the species previously placed into Tabulipora and Dyscritella could prove their re-assignment to Nikiforopora and Iraidina, respectively. Further detailed studies of bryozoans from the Lower Permian of Australia are needed in order to improve their use for stratigraphy and palaeobiogeography.
CONCLUSIONS
Three new species from the Callytharra Formation are described: Nikiforopora haigi sp. nov., Iraidina dendroidea sp. nov., and I. multicava sp. nov. The genera Nikiforopora and Iraidina are reported from the Permian deposits of Australia for the first time. Their importance for stratigraphy and palaeobiogeography is still restricted. Further intensive study of bryozoans from the Lower Permian of Australia are necessary.
ACKNOWLEDGEMENTS
David Haig, Arthur Mory, and Eckart Håkansson (all Perth) are thanked for organising the field work and their help in the field, as well as for providing additional bryozoan material from the Lower Permian of Western Australia. Deutsche Forschungsgemeinschaft (DFG) generously provided financial support which is appreciated (DFG project ER 278/7-1 and 2). Catherine Reid, Christchurch, and Patrick Wyse Jackson, Dublin, are thanked for their constructive reviews.